Introduction
De shikimate route is nauw verbonden met die van de aromatische aminozuren (L-tryptofaan, L-fenylalanine, en L-tyrosine) en vertoont in landplanten zeer hoge fluxen met schattingen van de hoeveelheid gefixeerde koolstof die door de route gaat variërend tussen 20 en 50% (Weiss, 1986; Corea et al., 2012; Maeda en Dudareva, 2012). Er is veel onderzoek verricht naar deze route omdat de aromatische aminozuren niet worden geproduceerd door mensen en monogastrisch vee en daarom een belangrijk voedingsbestanddeel zijn (Tzin en Galili, 2010). Bovendien is een van de enzymen van de pathway – 5-enolpyruvalshikimaat-3-fosfaat synthase (EPSP) – een van de meest gebruikte doelwitten voor herbiciden (zie, Duke en Powles, 2008). Bovendien worden, zoals wij onlangs hebben beschreven, plantaardige fenolische secundaire metabolieten en hun precursoren gesynthetiseerd via de weg van de shikimaatbiosynthese en de talrijke vertakkingen daarvan (Tohge et al., 2013). De shikimaatroute is in hoge mate geconserveerd en wordt aangetroffen in schimmels, bacteriën en plantensoorten, waar hij actief is in de biosynthese van niet alleen de drie hierboven beschreven aromatische aminozuren, maar ook van ontelbare aromatische secundaire metabolieten zoals alkaloïden, flavonoïden, ligninen, en aromatische antibiotica. Veel van deze verbindingen zijn bioactief en spelen een belangrijke rol in de verdediging van planten tegen biotische en abiotische stress en milieu-interacties (Hamberger et al., 2006; Maeda en Dudareva, 2012), en zijn als zodanig van groot fysiologisch belang. Geschat wordt dat onder normale omstandigheden maar liefst 20% van de totale vastgelegde koolstof door de shikimaatroute stroomt (Ni et al., 1996), met een grotere koolstofstroom door de route in tijden van stress of snelle groei van de plant (Corea et al., 2012). Gezien het belang ervan is het misschien niet verrassend dat alle leden van biosynthesegenen en overeenkomstige enzymen die betrokken zijn bij de shikimaatroute, zijn gekarakteriseerd in modelplanten zoals Arabidopsis. Vergelijking tussen soorten van de shikimaatbiosynthesenzymen heeft aangetoond dat ze sequentieovereenkomst, divergente evolutie en gemeenschappelijke reactiemechanismen vertonen (Dosselaere en Vanderleyden, 2001). Alle andere soorten verschillen echter aanzienlijk van schimmels, die een complex systeem hebben ontwikkeld met één enkel pentafunctioneel polypeptide, bekend als het AroM complex, dat vijf opeenvolgende reacties uitvoert (Lumsden en Coggins, 1977; Duncan et al., 1987). In dit overzicht geven wij een samenvatting van de huidige kennis betreffende de genetische aard van deze route, waarbij wij ons concentreren op vergelijkingen tussen verschillende soorten, die een breed spectrum van soorten omvatten, waaronder algen (Chlamydomonas reinhardtii, Volvox carteri, Micromonas sp, Ostreococcus tauri, Ostreococcus lucimarinus), mos (Selaginella moellendorffii, Physcomitrella patens), monocots (Sorghum bicolor, Zea mays, Brachypodium distachyon, Oryza sativa ssp. japonica en Oryza sativa ssp. indica), en dicots (Vitis vinifera, Theobroma cacao, Carica papaya, Arabidopsis thaliana, Arabidopsis lyrata, Populus trichocarpa, Ricinus communis, Manihot esculenta, Malus domestica, Fragaria vesca, Glycine max, Lotus japonicus, Medicago truncatula) soorten (tabel 1). Tenslotte vergelijken en contrasteren we de evolutie van deze pathway met die van de meer gespecialiseerde pathways van fenylpropanoïde biosynthese.
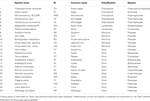
Tabel 1.
Shikimate Biosynthesis and Phenylalanine Derived Secondary Metabolism in Plants
Gezien het feit dat fenolische secundaire metabolieten die via shikimate biosynthese van fenylalanine worden afgeleid, wijd verspreid zijn in planten en andere eukaryoten, zijn de genen die coderen voor shikimate biosynthetische enzymen over het algemeen sterk geconserveerd in de natuur. Acht en twee reacties zijn betrokken bij respectievelijk de biosynthese van shikimaat en fenylalanine. Zowel de leden van alle genfamilies als de overeenkomstige biosynthetische enzymen die bij deze routes betrokken zijn, zijn gekarakteriseerd in modelplanten zoals Arabidopsis (Figuur 1A). Daarentegen vertonen fenolische secundaire metabolieten afgeleid van fenylalanine een aanzienlijke soortspecifieke distributie waarbij de fenolische secundaire metabolieten in het plantenrijk zoals cumarinederivaten, monolignine, lignine, spermidinederivaten, flavonoïde, tannine aanwezig zijn in specifieke families binnen de groene lineage (Figuur 1B). Deze diversiteit is ontstaan door de actie van diverse evolutionaire strategieën, bijvoorbeeld genverdubbeling en cis-regulerende evolutie om zich aan te passen aan de heersende milieuomstandigheden. Gezien hun soortspecifieke verspreiding worden de genen die betrokken zijn bij het fenolisch secundair metabolisme in planten, zoals fenylammonia-lyase (PAL), polyketidesynthase (PKS), 2-oxoglutaraat-afhankelijke deoxygenasen (2ODD’s) en UDP-glycosyltransferasen (UGT’s), vaak gebruikt als casestudies van de evolutie van planten (Tohge et al., 2013). Ondanks het feit dat shikimaat-fenylalanine biosynthesegenen goed geconserveerd zijn in alle soorten, inclusief algensoorten, werden fenol secundair metabolisme gerelateerde orthologe genen niet gedetecteerd in alle algensoorten (Tabel 2, Tohge et al., 2013). Dit resultaat suggereert een aanzienlijk oudere oorsprong van de shikimate-phenylalanine pathways. In de volgende paragrafen zullen we de evolutie van de shikimaat-phenylalanine-banen bespreken, met de nadruk op vergelijkingen tussen soorten voor elk gen dat codeert voor een van de samenstellende enzymen van een van beide banen.
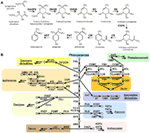
Figuur 1. De biosynthese van shikimaat en fenylalanine afgeleide secundaire metabolieten bij planten. (A) Shikimaatbiosynthese uitgaande van fosfoenolpyruvaat (PEP) en D-erytrose 4-fosfaat wordt beschreven met gekarakteriseerde genen en gerapporteerde tussenmetabolieten. (B) Fenylalanine afgeleide belangrijke fenolische secundaire meboliet biosynthese in de groene lineage. Pijl geeft enzymatische reactie aan, cirkel geeft metaboliet aan. Afkorting: DAHPS, 3-deoxy-D-arabino-heptulosonaat 7-fosfaat synthase; DQS, 3-dehydroquinaat synthase; DHQD/SD, 3-dehydroquinaat dehydratase; SK, shikimaat kinase; ESPS, 3-fososhikimaat 1-carboxyvinyltransferase; CS, chorismaat synthase; CM, chorismaat mutase; PAT, prefenaat aminotransferase; ADT, arogenaat dehydratase. PAL, fenylalanine-ammoniaklyase; C4H, cinnamaat-4-hydroxylase; 4CL, 4-coumaraat-CoA-ligase; CAD, cinnamoyl-alcoholdehydrogenase; F5H, ferulaat-5-hydroxylase; C3H, coumaraat-3-hydroxylase; ALDH, aldehyde-dehydrogenase; CCR, cinnamoyl-CoA-reductase; HCT, hydroxycinnamoyl-Coenzym A shikimaat/chinaat hydroxycinnamoyltransferase; CCoAOMT, caffeoyl/CoA-3-O-metheltransferase; CHS, chalcone synthese; CHI, chalcone isomerase; F3H, flavanon 3-hydroxylase; F3′H, flavonoid-3′-hydroxylase; F3GT, flavonoid-3-O-glycosyltransferase; FS, flavone synthase; FOMT, flavonoid O-methyltransferase; FCGT, flavone-C-glycosyltransferase; FLS, flavonol synthese; F3GT, flavonoid-3-O-glycosyltransferase; DFR, dihydroflavonolreductase; ANS, anthocyanidinesynthese; AGT, flavonoïd-O-glycosyltransferase; AAT, anthocyanine-cyltransferase; BAN, oxidoreductase|dihydroflavonolreductase-achtige; LAC, laccase.

Tabel 2. Shikimate en fenylalanine biosynthese genen en homologs in elke soort met/zonder tandem gedupliceerde genen.
3-Deoxy-D-arabino-heptulosonate 7-Phosphate Synthase
De eerste enzymatische stap van de shikimaatroute, 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase (DAHPS), katalyseert een aldolcondensatie van fosfoenolpyruvaat (PEP) en D-erytrose-4-fosfaat (E4P) om 3-deoxy-D-arabino-heptulosonaat-7-fosfaat (DAHP) te produceren (figuur 1). Op basis van hun eiwitstructuur kunnen DAHPS’s in twee verschillende homologieklassen worden ingedeeld. De van microben afgeleide klasse I DAHPS bevatten een bifunctioneel chorismaat mutase (CM)-DAHPS-domein, om die reden worden microbiële DAHPSs, bijvoorbeeld E. coli (AroF, G, en H) en S. cerevisiae (Aro3 en 4), geclassificeerd als klasse I DAHPSs. Daarentegen werd eerder gedacht dat DAHPS van klasse II alleen in plantensoorten voorkwamen, maar deze werden later gerapporteerd in bepaalde microben zoals Streptomyces coelicolor, Streptomyces rimosus en Neurospora crassa (Bentley, 1990; Maeda en Dudareva, 2012). De DAHPS (AroA) en CM (AroQ) activiteiten van B. subtilis DAHPS zijn echter gescheiden door domeintruncatie. Gedetailleerde sequentiestructuuranalyse van de bacteriële AroA- en AroQ-families, enzymatische studies met het volledige eiwit en de afgeknotte domeinen van AroA en AroQ van B. subtilis, en vergelijking met fusie-eiwitten van Porphyromonas gingivalis waarin het AroQ domein gefuseerd was met de C terminus van AroA, suggereren dat “feedback regulatie” inderdaad de evolutionaire link kan zijn tussen de twee klassen die geëvolueerd zijn uit primitieve ongereguleerde leden van klasse II DAHPS (Wu en Woodard, 2006). Klasse II plant DAHPSs zijn gerapporteerd van wortelwortels (Suzich et al., 1985) en aardappelcelcultuur (Pinto et al., 1986; Herrmann and Weaver, 1999). DAHPS wordt gecodeerd door drie genen in het Arabidopsis-genoom (AtDAHPS1, AT4G39980; AtDAHPS2, At4g33510; AtDAHPS3, At1g22410). Zoeken naar orthologe genen met behulp van de Arabidopsis DAHPSs, leverde één gen op in algensoorten (Chlamydomonas reinhardtii, Volvox carteri, Micromonas sp., en Ostreococcus tauri) en Lotus japonica, maar twee tot acht isovormen in andere hogere plantensoorten (Tabel 2). AtDAHPS1-type en AtDAHPS2-type genen vertonen differentiële expressie in Arabidopsis thaliana, Solanum lycopersicum, en Solanum tuberosum (Maeda en Dudareva, 2012). AtDAHPS1-type genen, die bovendien onderhevig zijn aan redoxregulatie door het ferredoxine-thioredoxine systeem, vertonen significante inductie door verwonding en pathogene infectie (Keith et al., 1991; Gorlach et al., 1995; Maeda en Dudareva, 2012), terwijl AtDAHPS2-type genen constitutieve expressie vertonen (Gorlach et al., 1995). Een fylogenetische analyse van de DAHPS-genen laat vier grote clades zien, (i) een microphyte-clade, (ii) een bryophyte-duplicatie-clade, (iii) een monocot en dicot houtachtige soorten-clade, (iv) een AtDAHPSs-clade (figuur 2Aa). Bovendien heeft hoofdcategorie iv vier subgroepen, (iv-a) AtDAHPS2-groep, (iv-b) monocot, (iv-c) AtDAHPS1-groep en (iv-d) AtDAHP3-groep. Dit resultaat geeft aan dat de constitutief tot expressie komende AtDAHPS1 en de stress responsieve AtDAHPS 3 type genen een goed geconserveerde sequentie vertonen tussen de soorten (clade iv-c en iv-d), terwijl de tweede constitutief tot expressie komende AtDAHPS2 type genen duidelijk gescheiden zijn tussen monocot en dicot soorten (clade iv-a).

Figuur 2. Fylogenetische boomanalyse van shikimaat- en fenylalaninebiosynthesegenen bij 25 soorten. Aminozuursequentie fylogenetische bomen van (A) shikimaatpathway: (a), DAHPS, (b) DHS, (c) DHQD/SD, (d) SK, (e) ESPS, en (f) CS, (B) fenylalanine-gerelateerde genen, (a) CM en (b) PAT. Aminozuursequenties van shikimate biosynthetische genen zijn verkregen uit de Plaza database (http://bioinformatics.psb.ugent.be/plaza/). Relaties tussen de beschouwde soorten worden gepresenteerd op de Plaza website. De fylogenetische boom werd geconstrueerd met de uitgelijnde eiwitsequenties door MEGA (versie 5.10; http://www.megasoftware.net/; Kumar et al., 2004) met behulp van de neighbor-joining methode met de volgende parameters: Poisson correctie, volledige schrapping, en bootstrap (1000 replicaten, willekeurig zaad). De eiwitsequenties werden uitgelijnd door Plaza. De waarden op de takken geven de bootstrap-ondersteuning in percentages aan.
3-Dehydroquinate Synthase
De tweede stap van de shikimate pathway wordt gekatalyseerd door 3-dehydroquinate synthase (DHQS), een enzym dat de intramoleculaire uitwisseling van de DAHP ringzuurstof met koolstof 7 bevordert om DAHP om te zetten in 3-dehydroquinate. In tegenstelling tot de hierboven beschreven schimmelsituatie is het DHQS-gen van de plant monofunctioneel en wordt het slechts als één enkele kopie aangetroffen in alle soorten, met uitzondering van Glycine max, dat twee genen in zijn genoom herbergt (figuur 2Ab). Fylogenetische analyse van DHQS-genen onthult drie grote clades bestaande uit (i) microphyte, (ii) bryophyte, (iii) monocot, (iv) Brassicaceae, en (v) dicot soorten. Intrigerend is dat de genexpressie van het DHQS-gen, in tegenstelling tot die van andere shikimate biosynthesegenen, niet goed gecorreleerd is met de productie van fenylpropanoïden in Arabidopsis (Hamberger et al, 2006).
3-Dehydroquinate Dehydratase/Shikimate Dehydrogenase
3-Deoxy-D-arabino-heptulosonate 7-phosphate wordt omgezet in 3-dehydroquinate door het bifunctionele enzym 3-dehydroquinate dehydratase/shikimate dehydrogenase (DHQD/SD), dat eerst de dehydratie van DAHP tot 3-dehydroshikimaat katalyseert en vervolgens de omkeerbare reductie van dit tussenproduct tot shikimaat, waarbij NADPH als co-factor wordt gebruikt. DHQD/SD bestaat in drie vormen; bacteriële specifieke klasse I shikimaatdehydrogenasen (AroE-type), klasse II shikimaat/quinaatdehydrogenasen (YdiB-type), en klasse III shikimaatdehydrogenase-achtige (SHD-L-type) (Michel et al., 2003; Singh et al., 2005). Bij planten van klasse IV is de enzymatische activiteit van DHQD 10 keer hoger dan die van SD, wat aangeeft dat de hoeveelheid 3-dehydroshikimaat meer dan voldoende is om de flux door de shikimaatroute te ondersteunen (Fiedler en Schultz, 1985). Dit bifunctionele enzym speelt een belangrijke rol bij de regulering van het metabolisme van verschillende fenolische secundaire metabolische routes (Bentley, 1990; Ding et al., 2007). In het algemeen bevatten zaadplanten één DHQD/SD-gen dat een sequentie bevat die codeert voor een plastisch transitpeptide in hun genoom (Maeda et al., 2011, Tabel 2). Een uitzondering op deze verklaring is Nicotiana tabacum die twee genen in zijn genoom bevat. Intrigerend is dat silencing van NtDHD/SHD-1 resulteert in sterke groeiremming en verlaging van het gehalte aan aromatische aminozuren, chlorogeenzuur en lignine (Ding et al., 2007), maar een tweede cytosolische isovorm kan de productie van shikimaat compenseren, maar niet op fenotypisch niveau. Meer algemeen blijkt uit de fylogenetische analyse dat microfyten ook een laag aantal DHQD/SD-genen bevatten (tussen één en twee), terwijl een duidelijke scheiding kon worden waargenomen tussen (i) de microfytenclade, (ii) de bryofytenclade, (iii) de monocotenclade, (iv) de houtachtige soortspecifieke tandem-genduplicatieclade, en (v) de dicotenclades (figuur 2Ac; tabel 2). Interessant is dat de observatie van de houtachtige soorten-specifieke tandem gen duplicatie clade suggereert dat deze soorten geëvolueerd zijn na DHQD/SD gen duplicatie. De cytosolische lokalisatie van NtDHD/SHD-2 is intrigerend omdat de aanwezigheid van DAHP synthase, ESPS synthase en CM isovormen zonder N-terminale plastid targeting sequenties is gerapporteerd (d’Amato, 1984; Mousdale en Coggins, 1985; Ganson et al., 1986). Bovendien suggereert de bevinding dat zowel ESPS synthase als shikimaat kinase (SK) actief zijn, zelfs wanneer zij hun doelsequenties behouden (Dellacioppa et al., 1986; Schmid et al., 1992), dat zij mogelijkerwijs ook bestanddelen van een cytosolische route zouden kunnen zijn. Tenslotte, experimenten waarbij geïsoleerde en zeer zuivere mitochondriën werden voorzien van 13C gelabelde glucose om de binding van de cytosolische isovormen van de glycolyse te onderzoeken (Giege et al., 2003) bracht ook 13C-verrijking in shikimaat aan het licht (Sweetlove en Fernie, 2013), wat erop wijst dat ook in deze soort waarschijnlijk een volledige cytosolische pathway bestaat.
Shikimate Kinase
De vijfde reactie van de shikimaat pathway wordt gekatalyseerd door SK, dat de ATP-afhankelijke fosforylering van shikimaat tot shikimaat-3-fosfaat (S3P) katalyseert. E. coli heeft twee SK’s, een van klasse I (AroL-type) en een van klasse II (AroK-type) die slechts 30% van de sequentie gemeen hebben (Griffin en Gasson, 1995; Whipp en Pittard, 1995; Herrmann en Weaver, 1999). In planten worden verschillende aantallen SK isovormen gevonden in verschillende soorten; slechts één in groenwieren, lycofyten en bryofyten, maar tussen één en drie in monocot en dicot planten (tabel 2). Een fylogenetische analyse van SK genen laat vijf grote clades zien, bestaande uit (i) microphyte, (ii) bryophyte, (iii) dicot houtige soort-specifieke clade, (iv) monocot clade, en (v) dicot soorten clade (Figuur 2Ad). Analyses van het SK-eiwit van de Spinacia olerancea hebben aangetoond dat het gemoduleerd wordt door de energiestatus en dus vergelijkbaar is met het SK-eiwit van bacteriën en andere ATP-utiliserende enzymen (Pacold en Anderson, 1973; Huang et al., 1975; Schmidt et al., 1990). Om deze reden is recent gepostuleerd dat SK een link zou kunnen leggen tussen de energievragende shikimate pathway en de cellulaire energiebalans (Maeda en Dudareva, 2012), maar directe experimentele ondersteuning voor deze hypothese ontbreekt op dit moment. In Arabidopsis is aangetoond dat homologe genen, SKL1 en SKL2 genaamd, die functioneel vereist zijn voor chloroplast biogenese, ontstaan zijn uit SK gen duplicatie (Fucile et al., 2008). SKL1 en SKL2 orthologs zijn gevonden in verschillende zaadplant soorten, maar niet in groene algen (tabel 2).
5-enolypyruvylshikimaat-3-fosfaat synthase
Het 5-enolypyruvylshikimaat-3-fosfaat synthase (EPSPS, 3-phosphoshikimate 1-carboxyvintltransferase) is de zesde stap en hier wordt een tweede PEP gecondenseerd met S3P om 5-enolypyruvylshiukimaat-3-fosfaat (EPSP) te vormen. Aangezien EPSPS het enige bekende doelwit is voor het herbicide glyfosaat (Steinrucken en Amrhein, 1980), worden de isovormen van dit enzym vaak geclassificeerd volgens hun gevoeligheid voor glyfosaat; glyfosaatgevoelige EPSPS klasse I komt voor in bacteriën en plantensoorten, terwijl glyfosaatongevoelige EPSPS klasse II is gerapporteerd in bepaalde bacteriën zoals Agrobacterium (Fucile et al., 2011). In planten wordt een verschillend aantal EPSPS isovormen gevonden in verschillende soorten; slechts één isovorm in groenwieren, lycofyten en bryofyten, maar één of twee worden gevonden in monocot en dicot soorten (Tabel 2). Fylogenetische analyse van EPSPS-genen toonde aan, atypisch voor genen geassocieerd met shikimate metabolisme, dat vijf grote groepen konden worden waargenomen; (i) microphyte, (ii) bryophyte, (iii) Brassicaceae specifieke clade, (iv) monocot soorten, en (v) dicot soorten clade (Figuur 2Ae). Er zijn duidelijke aanwijzingen dat gedupliceerde EPSPS-genen in Arabidopsis, appel, wijnstok, sojaboon en populier het resultaat zijn van onafhankelijke duplicatiegebeurtenissen binnen hun lineages, waarbij beide kopieën in Arabidopsis behouden blijven (Hamberger et al, 2006), maar de reden voor de unieke divergentie in dit gen van de pathway is momenteel onduidelijk.
Chorismate Synthase
Chorismate, het eindproduct van de shikimate pathway, wordt vervolgens gevormd door chorismate synthase (CS) dat de trans-1,4 eliminatie van fosfaat uit EPSP katalyseert. CS worden ingedeeld in twee functionele groepen (i) schimmel type bifunctionele CS die geassocieerd zijn met NADPH-afhankelijk flavin reductase of (ii) bacteriële en plant type monofunctionele CS (Schaller et al., 1991; Maeda and Dudareva, 2012). Voor de door CS gekatalyseerde reactie is flavin mononucleotide (FMN) nodig en de totale reactie is redoxneutraal (Ramjee et al., 1991; Macheroux et al., 1999; Maclean and Ali, 2003). FMN is een elektronendonor voor EPSP die de splitsing van fosfaat vergemakkelijkt. Het eerste gekloonde CS-gen voor planten was dat van C. sempervirens (Schaller et al., 1991), dat een enkel CS in zijn genoom bevat. Aangezien dit gen een 5′ plastide import signaalsequentie heeft, wijzen deze resultaten erop dat er bij deze soort wellicht geen CS buiten de plastide bestaat. Onderzoek van andere soorten bracht aan het licht dat één tot twee CS-genen aanwezig waren in groenwieren, lycofyten, en bryofyten, alsook in dicot soorten, maar dat één tot drie genen aanwezig zijn in de genomen van appel- en peulvruchtensoorten (Tabel 2). Een fylogenetische analyse van CS genen onthult drie belangrijke clades gevormd door (i) microphyte, (ii) monocot, (iii) dicot soorten (Figuur 2Af).
Chorismate Mutase
Chorismate mutase katalyseert de eerste stap van fenylalanine en tyrosine biosynthese en vertegenwoordigt bovendien een belangrijke stap in de richting van de vertakking van tryptofaan biosynthese. CM katalyseert de omzetting van chorismaat in prefenaat via een Claisen-herschikking. De bacteriële minor CM proteïnen (AroQ type, klasse I CM) vertonen monofunctionele enzymatische activiteit, terwijl verschillende bifunctionele CMs zoals CM-PDT, CM-PDH, en CM-DAHP daarnaast zijn gevonden in schimmels en bacteriën (klasse II CM, Euverink et al., 1995; Romero et al., 1995; Chen et al., 2003; Baez-Viveros et al., 2004). Ondanks het feit dat er slechts één CM gen aanwezig is in de genomen van algen en lycofyten, worden er meer dan één genkopie (twee tot vijf) gevonden in zowel bryofyten als monocot en dicot soorten (Tabel 2). In zaadplanten draagt CM1 een putatief plastid transit peptide, maar CM2 niet en is bovendien meestal ongevoelig voor allosterische regulatie door aromatische aminozuren (Benesova en Bode, 1992; Eberhard et al., 1996; Maeda en Dudareva, 2012). Verschillende plantensoorten, vooral dicot planten, hebben een extra CM3 familiegen dat een hoge sequentie gelijkenis vertoont met CM2 maar een vermoedelijke plastid transit peptide draagt. Arabidopsis heeft bijvoorbeeld drie isozymes, AtCM1 (At3g29200), AtCM2 (At5g10870) en AtCM3 (At1g69370) (Mobley et al., 1999; Tzin and Galili, 2010). Fylogenetische analyse van de CS-genen onthult drie grote clades die bestaan uit (i) AtCM2-clade, (ii) microfyten- en bryofyten-clade, en (iii) AtCM2-clade (figuur 2Ba). Bovendien vertoont clade iii twee subgroepen, (iii-a) AtCM3-subgroep en (iii-b) AtCM1-subgroep (figuur 2Ba) (Eberhard et al., 1996). Ondanks dat de CM2 subgroep alle soorten zaadplanten bevat, zijn monocot soorten niet opgenomen in AtCM3 subgroep. Recent is het belang van CM uitgebreid tot buiten het intracellulaire metabolisme. In Zea mays is het chorismaat mutase Cmu1, uitgescheiden door Ustilago maydis, een wijdverspreide ziekteverwekker die gekenmerkt wordt door de ontwikkeling van grote plantentumoren en algemeen bekend staat als smut, een virulentiefactor. De opname van het Ustilago CMu1-eiwit door plantencellen maakt een omleiding van het metabolisme van de plant mogelijk en verandert de metabolische status van deze cellen via metabolic priming (Djamei et al., 2011). Het blijkt nu dat gesecreteerde CM’s worden aangetroffen in veel plantgerelateerde microben en deze vorm van gastheermanipulatie lijkt een algemeen wapen in het arsenaal van plantpathogenen.
Prefenaataminotransferase en Arogenaatdehydratase
Prefenaataminotransferase (PAT) en arogenaatdehydratase (ADT) katalyseren de laatste stappen voor de productie van fenylalanine. Terwijl ADT voor het eerst werd gekloond in 2007 (Cho et al., 2007; Huang et al., 2010), is het pas recenter dat PAT werd gekloond. In 2011 gepubliceerde artikelen identificeerden PAT in Petunia hybride, Arabidopsis thaliana en Solanum lycopersicum (Dal Cin et al., 2011; Maeda et al., 2011) en stelden vast dat het de koolstofflux van prefenaat naar arogenaat leidt, maar ook dat het sterk en gecoördineerd wordt geüpreguleerd met genen van het primaire metabolisme en van fenylalanine afgeleide smaakvervluchtingen. In plantensoorten is een verschillend aantal PAT-isovormen gevonden. Hoewel groene algen slechts één PAT en ADT genen bevatten, hebben monocot soorten tussen één en twee PATs en tussen twee en vier ADTs, terwijl dicot planten genomen hetzelfde aantal PATs bevatten maar twee tot acht ADTs (tabel 2). Fylogenetische analyse van PAT-genen toont drie grote clades van (i) microphyt, (ii) monocot, en (iii) dicot soorten (Figuur 2Bb).
Gen Betrokken bij Plant Phenolic Secondary Metabolisms
Phenolic secondary metabolism vertoont een immense chemische diversiteit als gevolg van de evolutie van enzymatische genen die betrokken zijn bij de verschillende biosynthetische en decoratieve pathways. Deze variatie wordt veroorzaakt door de diversiteit en redundantie van verscheidene sleutelgenen van het fenolisch secundair metabolisme, zoals PKS’en, cytochroom P450’s (CYP’s), Fe2+/2-oxoglutaraat-afhankelijke dioxygenasen (2ODD’s) en UDP-glycosyltransferasen (UGT’s). Anderzijds zijn er andere algemene aan fenylpropanoïden gerelateerde biosynthesegenen, fenylalanine-ammoniak-lyase (PAL), cinnamaat-4-hydroxylase (C4H) en 4-coumaraat:co-enzym-A-ligase (4CL), die nodig zijn voor de differentiatie van verschillende klassen van het fenolisch secundair metabolisme. Al deze kerngenen coderen voor belangrijke enzymen die een aantal hydroxycinnamzuren activeren om precursoren te leveren voor de biosynthese van ligninen, monolignalen, en inderdaad alle andere belangrijke fenolische secundaire metabolieten in hogere planten (Lozoya et al., 1988; Allina et al., 1998; Hu et al., 1998; Ehlting et al., 1999; Lindermayr et al., 2002; Hamberger en Hahlbrock, 2004). Aangezien het fenolische secundaire metabolisme een aanzienlijke soortspecificiteit vertoont, wordt het onderzoek van de genen die coderen voor de verantwoordelijke biosynthetische enzymen vaak gebruikt als een voorbeeld van chemotaxonomie om de evolutie van planten te begrijpen. De evolutie van deze geïsoleerde genen is echter eerder beperkend en een dieper inzicht wordt verschaft door dit te combineren met onderzoek van de evolutie van de shikimaat-fenylalanine biosynthesegenen in de groene lineage.
Conclusie
Tijdens de lange evolutionaire periode die zich uitstrekt van aquatische algen tot landplanten, hebben planten zich aangepast aan de niches van het milieu met evolutionaire strategieën zoals genverdubbeling en convergente evolutie door de filtratie van natuurlijke selectie. Genen van de plant shikimaat biosynthese zijn dienovereenkomstig geëvolueerd (figuur 3). In dit overzicht hebben we aangetoond dat de biosynthesegenen van het primaire metabolisme van aromatische aminozuren goed geconserveerd zijn tussen algen en alle landplanten. Echter, in tegenstelling tot algensoorten die noch isovormen noch gedupliceerde genen in hun genomen hebben, herbergen alle landplanten gen duplicaties inclusief tandem gen duplicaties die bijzonder prominent zijn in het geval van DAHPS, DHQD/SD, CS, CM, en ADT (Figuur 3A; Tabel 2). Onze fylogenetische analyse onthulde een duidelijke scheiding tussen algen, monocoten, dicoten, houtachtige soorten, en peulvruchten. Analyse van de aanwezigheid en het aantal kopieën van belangrijke genen in deze soorten geeft verschillende aanwijzingen over hoe we ons begrip van de steiger van waaruit deze genen zijn geëvolueerd kunnen verbeteren. De exacte evolutionaire druk op de genen van de shikimate biosynthese, met inbegrip van het unieke voorkomen van het Arom complex, zal echter nog heel wat verdere studies vergen. Dit gezegd zijnde, is het intrigerend om de biosynthesegenen te vergelijken en te contrasteren met de genen die stroomafwaarts van hen liggen in de productie van plantaardige fenolen (Figuur 3B). Interessant is dat de genen van de shikimate pathway alomtegenwoordig zijn in de groene lineage, terwijl dit niet kan worden gezegd van alle downstream genen van de fenylpropanoïde biosynthese. Bovendien is er een veel grotere gen duplicatie binnen phenylpropanoid dan shikimate biosynthese (figuur 3A; tabel 2). Dit feit komt ook tot uiting in het niveau van de chemische diversiteit van de respectieve routes, waarbij het essentiële karakter van de shikimaatroute veel diversiteit verhindert, maar de fenylpropanoïdesoorten in functie vaak overbodig zijn ten opzichte van elkaar. Het lijkt waarschijnlijk dat de fenylpropanoïdenroute aanvankelijk is ontstaan door mutaties die zich in de genen van de shikimatenroute ophoopten. Maar hoewel deze potentieel gunstig waren in landplanten om redenen die we bespreken in ons recente overzicht van deze verbindingen (Tohge et al., 2013), lijken ze de essentialiteit van shikimaat niet te delen in de hele groene lineage.
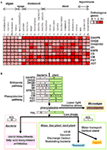
Figuur 3. Heat map voor isovormen van shikimate-phenylalanine biosynthesegenen in plantengenen en hypothetisch schema voor de evolutie van fenylalanine afgeleid fenolisch secundair metabolisme. (A) Stapelkaart overzicht van het aantal shikimaat-fenylalanine biosynthese gen-isovormen in 25 soorten. (B) Hypothetische schematische figuur voor shikimaat-phenylalanine biosynthese genen en hun evolutie van fenolisch secundair metabolisme.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
De onderzoeksactiviteit van Takayuki Tohge wordt ondersteund door de Alexander von Humboldt Foundation. Financiering door de Max-Planck-Society (aan Takayuki Tohge, Mutsumi Watanabe, Rainer Hoefgen, Alisdair R. Fernie) wordt dankbaar erkend.
Benesova, M., and Bode, R. (1992). Chorismate mutase isoforms from seeds and seedlings of Papaver somniferum. Phytochemistry 31, 2983-2987.
CrossRef Full Text
Fiedler, E., and Schultz, G. (1985). Localization, purification, and characterization of shikimate oxidoreductase-dehydroquinate hydrolase from stroma of spinach-chloroplasts. Plant Physiol. 79, 212-218.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hamberger, B., Ehlting, J., Barbazuk, B., and Douglas, C. J. (2006). Comparative genomics of the shikimate pathway in Arabidopsis, Populus trichocarpa and Oryza sativa: shikimate pathway gene family structure and identification of candidates for missing links in phenylalanine biosynthesis. Recent Adv. Phytochem. 40, 85-113.
CrossRef Full Text
Maclean, J., and Ali, S. (2003). De structuur van chorismate synthase onthult een nieuwe flavin binding aan een unieke chemische reactie. Structure 11, 1499-1511.
Pubmed Abstract | Pubmed Full Text | Pubmed Full Text | CrossRef Full Text
Schmid, J., Schaller, A., Leibinger, U., Boll, W., and Amrhein, N. (1992). De in vitro gesynthetiseerde tomaat shikimate kinase precursor is enzymatisch actief en wordt door chloroplasten geïmporteerd en verwerkt tot het rijpe enzym. Plant J. 2, 375-383.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tzin, V., and Galili, G. (2010). Nieuwe inzichten in de biosynthesewegen van shikimaat en aromatische aminozuren in planten. Mol. Plant 3, 956-972.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text